极化上皮细胞的蛋白质分类和转运
高等生物,如哺乳动物,由许多器官组成,它们都完成特定的任务,需要进行交流。为了实现定向的、可控的通信,这些功能单元必须在空间和生物化学上分离。因此,所有的动物器官都被上皮细胞包围着,这些上皮细胞除了保护它们免受有害环境的影响外,还确保它们与周围环境的物质交换受到控制。最小的单位是极化上皮细胞.这些细胞形成的屏障厚度正好是由一排不间断的细胞组成的细胞层。细胞之间的紧密连接创造了一个密封的表面,受控的运输过程通过这个表面发生。
上皮细胞通常具有极性结构。最显著的特征是细胞质膜的差异。有两个不同的域:顶端的面向器官外部的膜基底外侧面向器官内部或上皮内相邻细胞的膜。两个膜结构域表现出明显不同的蛋白质和脂质成分。例如,各种消化酶,如LPH(乳糖酶-根皮苷水解酶)或SI(蔗糖异麦芽糖酶)这些酶只运输到肠上皮的顶端细胞质膜,在那里它们到达它们的作用部位——肠腔。这些酶在基底外侧膜中找不到。将它们运输到那里没有意义——这将是一种能量浪费。
那么这种蛋白质分类的基本机制是什么呢?答案是由蛋白质本身提供的,它们都带有特异性topogenous信号。这些可以是短氨基酸序列(YXXφ, NPXY, LL, L),就像在基底外侧运输的蛋白质中发现的那样。信号效应也可能是基于蛋白质的糖基化,就像各种顶端蛋白的情况一样。GPI锚(糖基-磷脂酰肌醇)也作为顶端分选信号。所有的拓扑信号都是其他蛋白质或脂质结构的识别代码,然后充当分类受体或分类平台。例如,一些基底外侧蛋白通过在反高尔基网络中网格蛋白依赖转运的适配器蛋白(AP1A)进行分类并转运到目标膜[2](冈萨雷斯和罗德里格斯·博兰,2009年)。另一方面,具有GPI锚定的蛋白质与特定的富含胆固醇的脂质结构——脂筏——表现出高度的亲和力,脂筏是顶膜的分选平台。还有其他的顶端蛋白像P75-绿色荧光蛋白由于糖结合蛋白的特殊糖基化作用,它们被识别并结合半乳糖凝集素-3,最终导致转运到正确的膜域(图1)[1].
为了更详细地研究蛋白质被分类和运输到极化上皮细胞的顶端膜的过程,显而易见的步骤是在显微镜下仔细观察这个细胞极。蒂尔夫显微镜(全内反射荧光)对这类研究特别有用,因为它提供了顶端细胞质膜上、内和紧靠顶端细胞质膜下的结构的非常详细的图像。这适用于固定细胞和活细胞。
图1:极化上皮细胞中的蛋白质分选-极化上皮细胞具有两个不同的质膜域-顶端和基底外侧-它们的蛋白质和脂质组成不同,并由紧密连接分隔。一些顶端(X,Y)和基底外侧的蛋白质分选机制已经被发现。一种顶端通路是由碳水化合物结合蛋白半乳糖凝集素-3控制的,半乳糖凝集素-3与一些顶端分选蛋白如P75-相连绿色荧光蛋白、LPH或GP-114。这个分选过程发生在后高尔基腔室,最有可能是一个回收核内体或与它紧密相连。ER:内质网,TGN:反高尔基网络,PGC:后高尔基室。
TIRFM可视化顶端蛋白及其分选受体
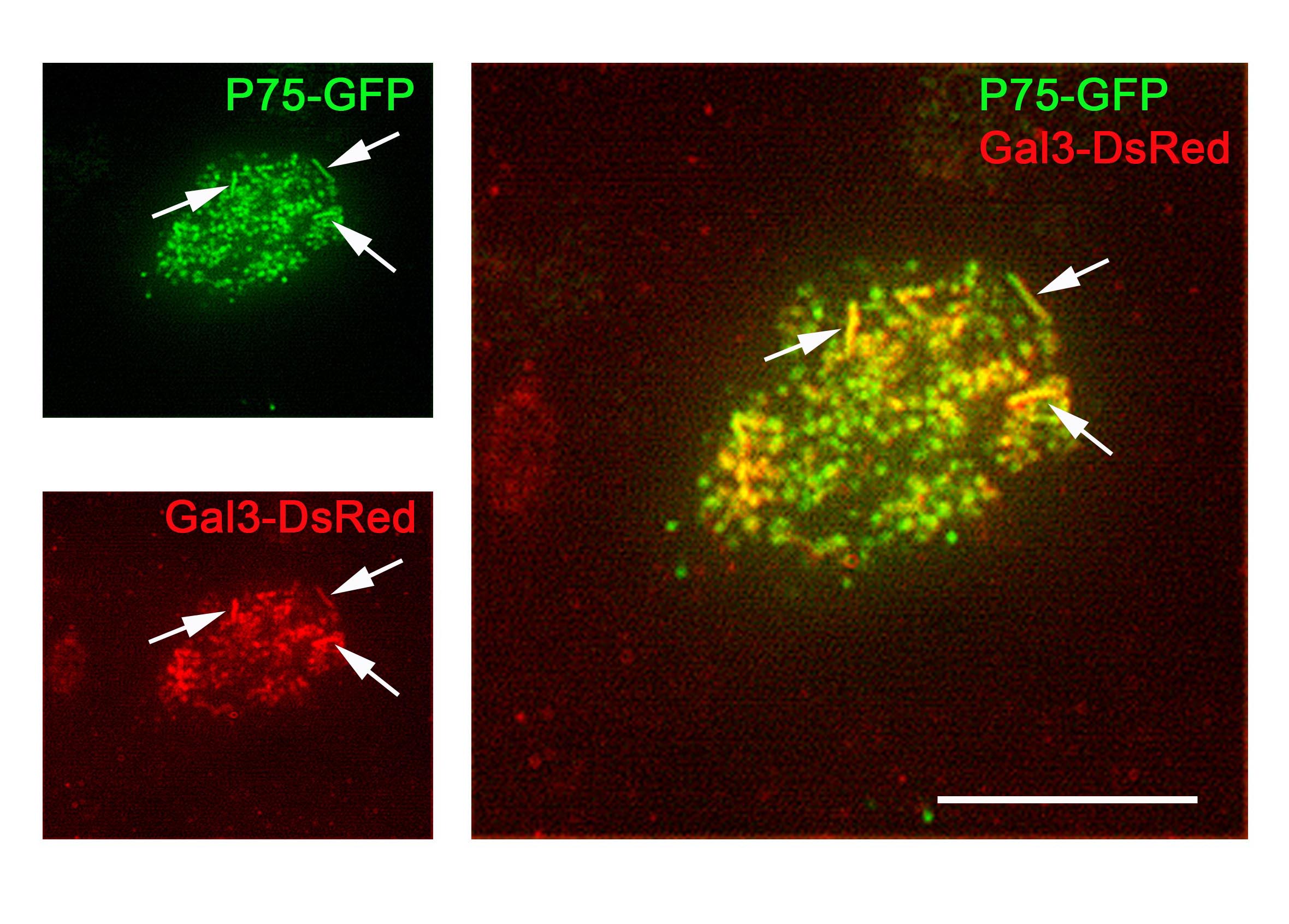
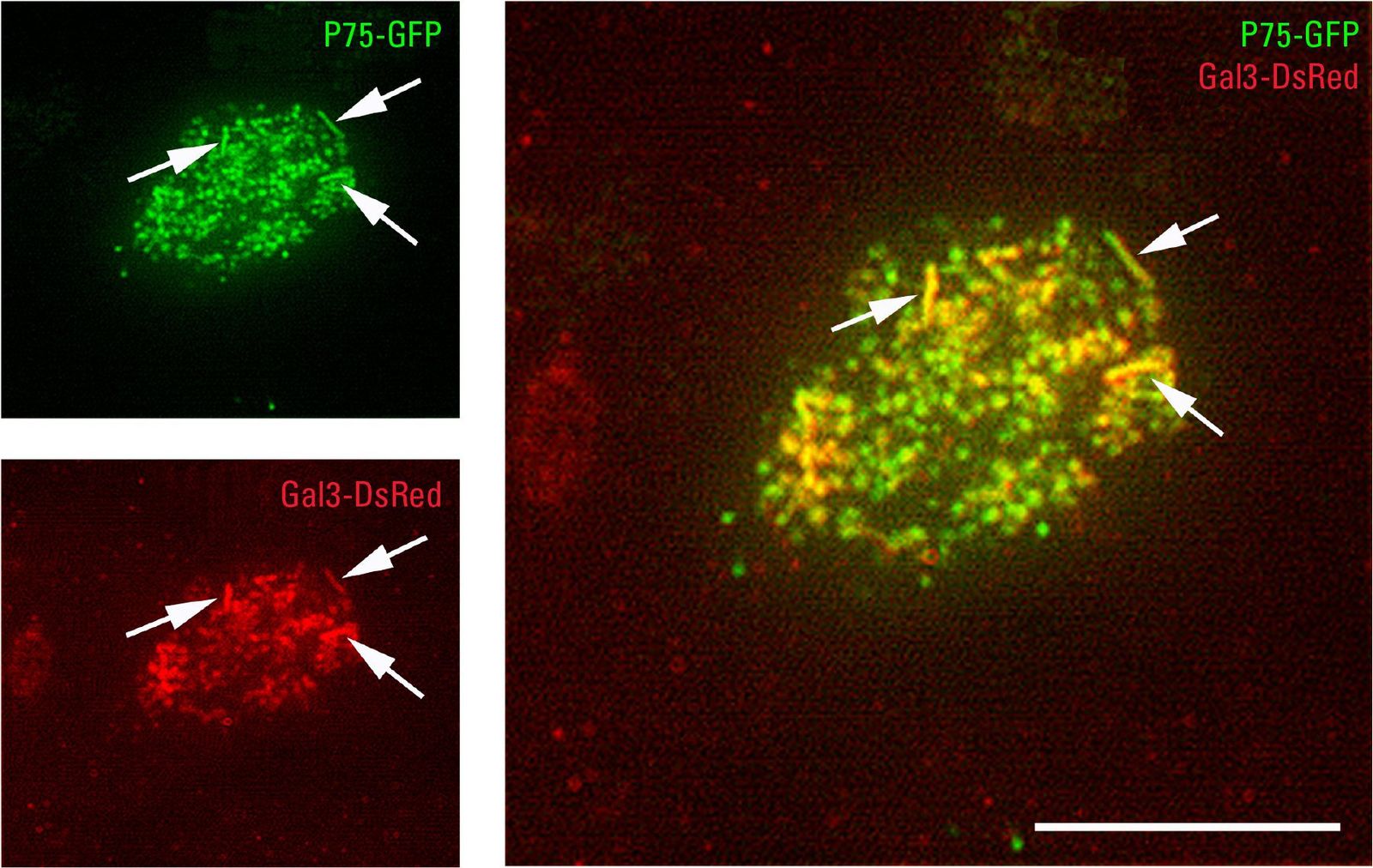
在显微镜下,用荧光蛋白标签标记被转运到顶膜或参与转运过程的蛋白质(例如:绿色荧光蛋白)或免疫荧光染色。根尖TIRFM是一种合适的方法,因为它能够分辨在其他显微镜技术中未检测到的根尖细胞极的结构[4].这意味着顶端蛋白(如P75-绿色荧光蛋白)可以与它们的分选受体半乳糖凝集素-3- dsred一起成像。共定位事件表明这两个蛋白之间存在相互作用(图2)。因此,TIRFM能够在体内可视化已被生化方法证实的顶端蛋白运输中的蛋白-蛋白相互作用。就像消失的场蒂尔夫显微镜穿透细胞的深度不超过300 nm,靠近细胞膜的区域清晰可见,使蛋白质相互作用位点位于顶膜内或正下方。
TIRFM在顶端细胞质膜上显示管状结构,很可能是微绒毛和细胞内小体结构。某些核内体被认为是蛋白质分拣中心。因此可以同时检测到顶端分选受体半凝集素-3及其靶蛋白P75-绿色荧光蛋白在rab11 -正循环核内体中[3.].
因此,TIRFM可以用来表明顶端蛋白P75-绿色荧光蛋白其分选受体半乳糖凝集素-3存在于亚根尖循环核内体中。这表明它可能是蛋白质的分选位点。
未来研究的有趣方法是借助TIRFM对蛋白质分类过程进行实时观察。由于大多数顶端蛋白质的强膜染色,因此有必要借助诱导蛋白质系统来跟踪顶端蛋白质从内质网到细胞质膜的路径预计起飞时间。
极化上皮细胞顶膜内吞过程的观察
已经提到的碳水化合物结合半乳糖凝集素-3在顶端蛋白分选过程中起作用。半乳糖凝集素-3被认为是被证实的顶端蛋白P75-的分拣受体绿色荧光蛋白和gp - 114。这种凝集素的下调导致P75-的基底外侧转运绿色荧光蛋白和GP-114[1].除了上面提到的寻找蛋白质分选过程的位点之外,还有一个问题是,一旦Galectin-3完成了它作为分选受体的任务后,它的命运如何。在生物化学上,已经证明一些凝集素从细胞传递到顶端培养基。在基底外侧培养基中几乎找不到半乳糖凝集素-3。不幸的是,用TIRFM还不能记录这种胞吐过程。
另一方面,半乳糖凝集素-3被极化的上皮细胞如MDCK细胞重新内吞。将重组alexa633偶联半乳糖凝集素-3应用于细胞的顶端,可以观察到这一过程。在TIRFM图像中,实验开始时细胞表面可见点状红色Galectin-3信号。随着记录的继续,这些斑点变得更弱、更扩散,因为被吸收的半乳糖凝集素-3- alexa633在被吸收后离开了TIRFM的消失场。如果在三维堆叠中观察到相同的实验(用表面荧光显微镜记录),就可以清楚地看到半乳糖凝集素-3- alexa633在实验过程中被摄取到细胞内间隔(可能是核内体)(图3)。
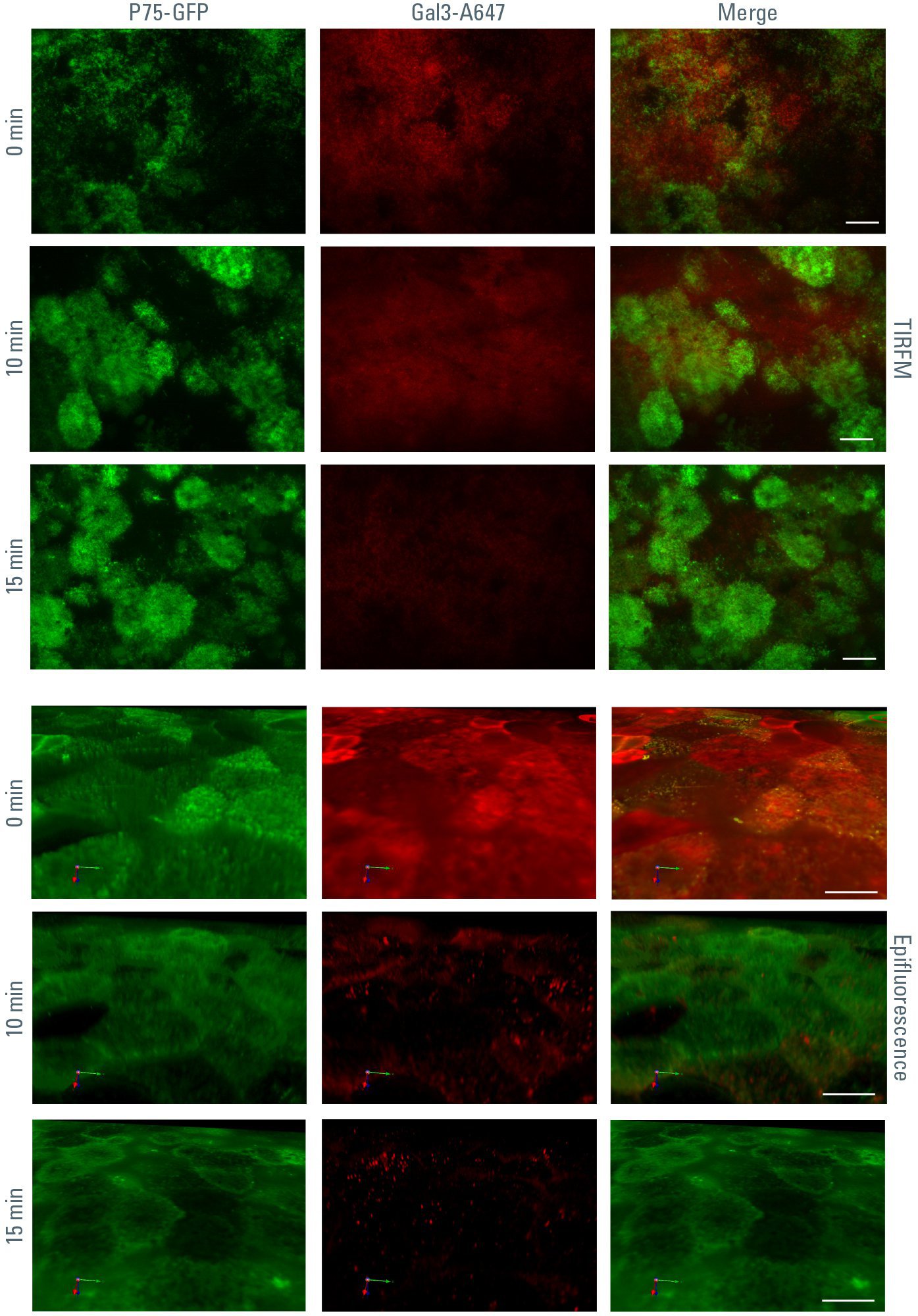
图3:重组半乳糖凝集素-3- a633在根尖细胞极被内吞-在37℃开始内吞后,半乳糖凝集素-3- a633根尖的TIRFM信号逐渐消失。从根尖侧观察到的三维荧光数据显示,半乳糖凝集素-3- a633信号从MDCK细胞根尖表面移动到细胞内内体结构。这些结构的位置可与P75-的质膜染色相比较绿色荧光蛋白.开始内吞作用后的时间在左边。渗透深度90 nm,垢棒10µm。
半乳糖凝集素-3、P75-GFP和顶端细胞骨架元件
如前所述,TIRFM改善了极化上皮细胞顶端表面单个结构的细节呈现。它不仅可以成像细胞内的隔间,还可以成像细胞质膜和相关蛋白质。这样,顶端细胞极的肌动蛋白细胞骨架可以通过phalloidin coup染色显示led荧光染色。结果是许多管状形状的塑料图像,除其他外,显示MDCK细胞中突出的微绒毛。顶端蛋白P75-绿色荧光蛋白(图4.1A)。然而,不是所有的P75-绿色荧光蛋白-标记区域位于微绒毛中(箭头)。很可能,这些是上述具有相似形状的循环内体。
如果在顶端TIRFM中观察到MDCK细胞带有某种形式的微管蛋白(乙酰化的微管蛋白),可以看到单一的长管状结构。这是初级纤毛(图4.1B),它是细胞的机械感觉工具。顶端蛋白P75-绿色荧光蛋白该蛋白及其分选受体半乳糖凝集素-3的囊泡转运可以沿着或在活体MDCK细胞的初级纤毛中观察到(图4.2)。这也可能是导致凝集素分泌的一个过程。
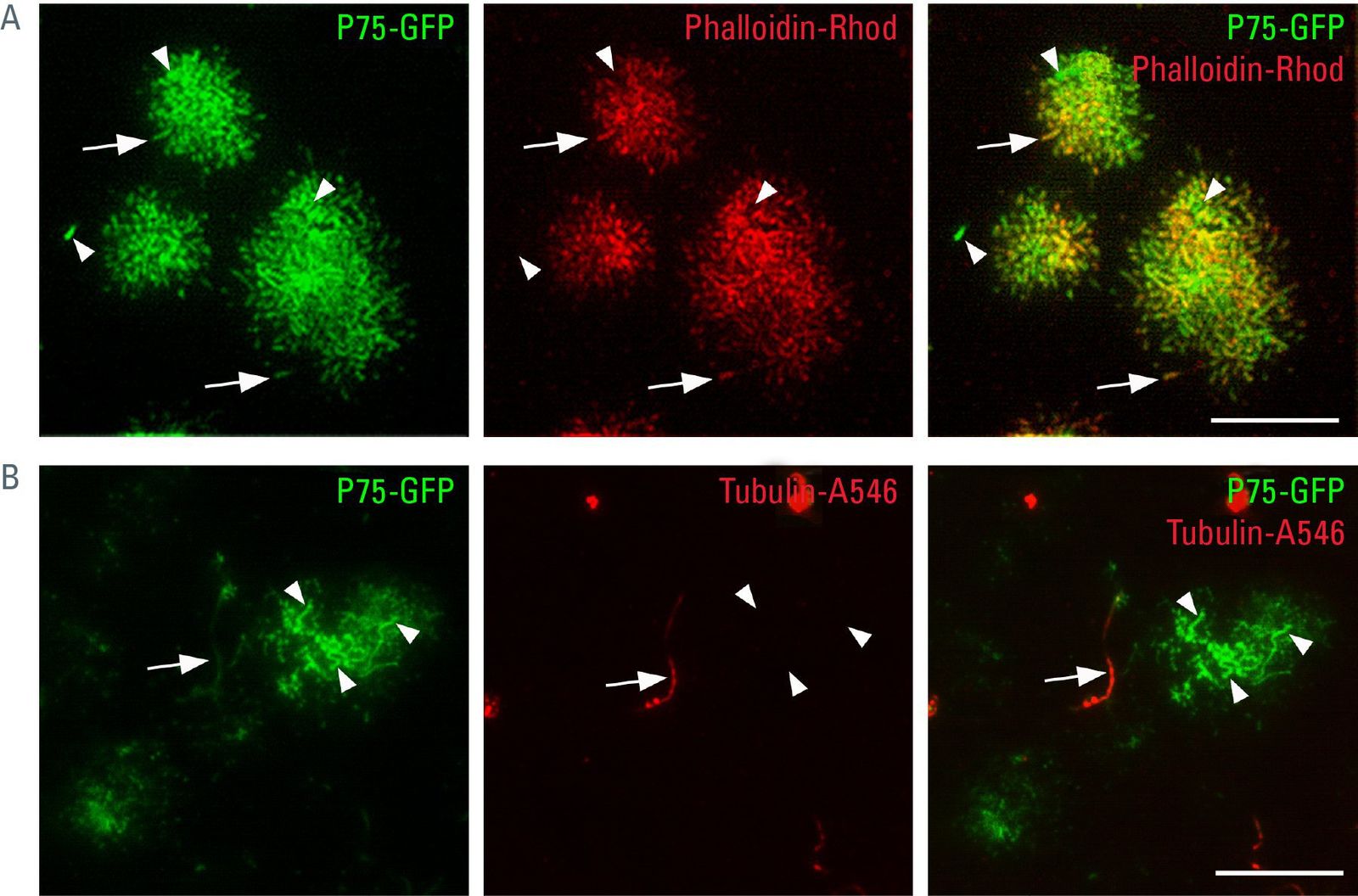
图4.1:我-绿色荧光蛋白与TIRFM - A细胞骨架结构相关)顶端蛋白P75-绿色荧光蛋白可在微绒毛结构中发现,微绒毛结构由肌动蛋白结合的鬼笔环素-罗丹明染色(箭头)。此外还有P75-绿色荧光蛋白微绒毛外可检测到信号,很可能是循环核内体(箭头)。B)我-绿色荧光蛋白不仅出现在微绒毛(箭头)中,也出现在初生纤毛(箭头)中,初生纤毛中特异性乙酰化微管蛋白染色,穿透深度90 nm,鳞片条10µm。
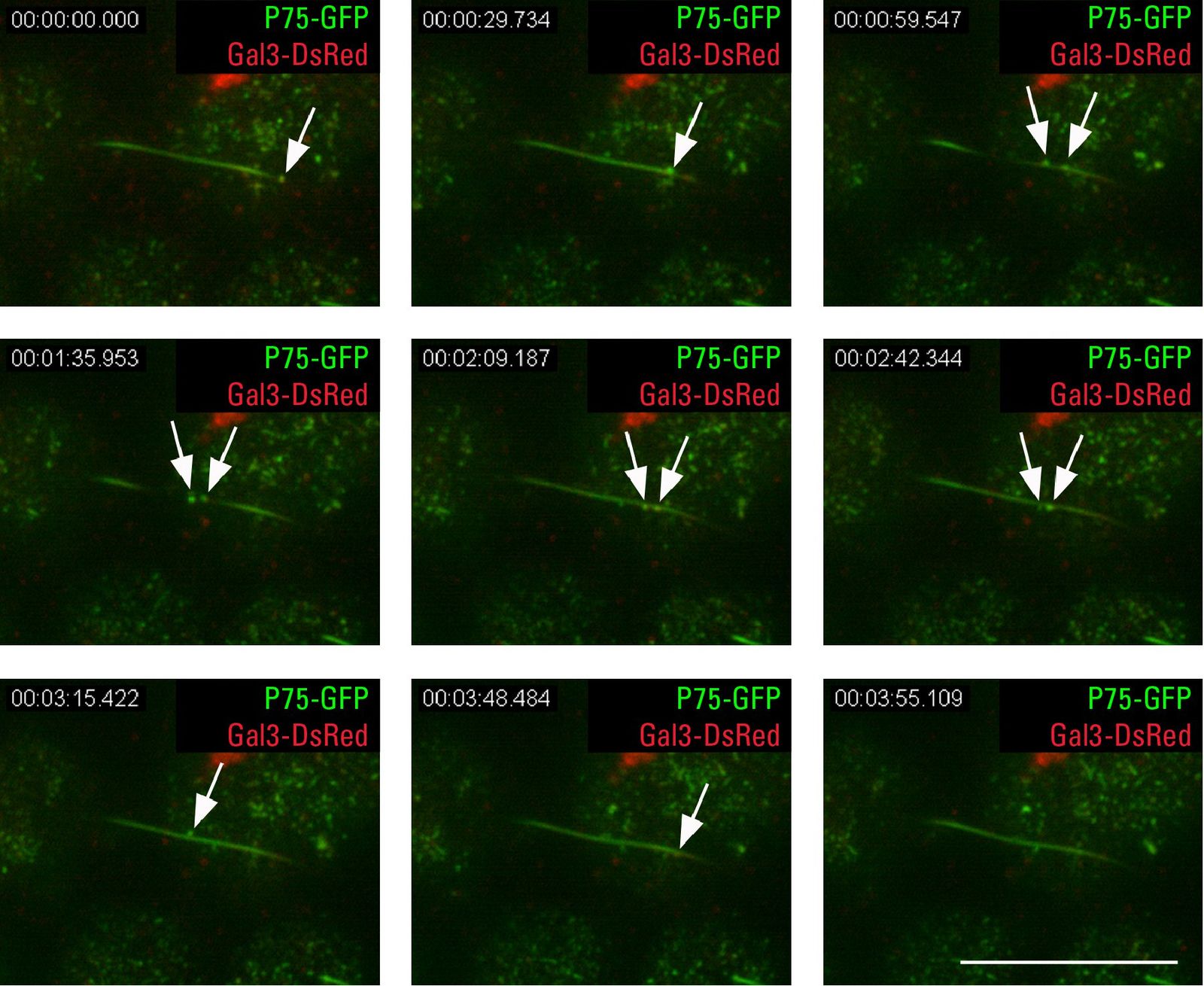
图4.2和视频:P75-绿色荧光蛋白和初生纤毛处半凝集素-3- dsred -顶端TIRFM生命细胞成像可与P75-一起破译半凝集素-3- dsred的泡状运动绿色荧光蛋白(箭头)沿着MDCK细胞初生纤毛。绝对时间3:55 min,渗透深度90 nm,垢条10µm。
结论
总之,可以说,TIRFM可以显著改善极化上皮细胞的顶端细胞极感兴趣区域的视野,在这里顶端分选蛋白如P75-绿色荧光蛋白可以和它们的分选受体半乳糖凝集素-3一起观察。也有可能看到潜在的分类隔间-回收核内体。TIRFM不仅显示了分泌途径上的蛋白质运输过程,还显示了半乳糖凝集素-3重新被细胞吸收的内吞过程。这些过程可以在固定细胞和活细胞中观察到。在后一种情况下,也有可能记录Galectin-3和P75-的转运事件绿色荧光蛋白在初生纤毛中。
参考文献
- Delacour D, Cramm-Behrens CI, Drobecq H, Le BA, Naim HY, Jacob R:半乳糖凝集素-3在顶端蛋白分类中的需求。咕咕叫生物16(2006)408 - 414。
- 网格蛋白和AP1B:通过跨内体途径的基底外侧转运的关键作用。FEBS Lett 583 (2009)3784 - 3795。
- Schneider D, Greb C, Koch A, Straube T, Elli A, Delacour D, Jacob R: Galectin-3通过极化和非极化细胞内体细胞器的转运。Eur J CellBiol (2010).http://www.ncbi.nlm.nih.gov/pubmed/20705359
- 格雷布C,雅各布R:荧光增强器和电极上皮细胞——TIRF Mikroskopie auf den Kopf gestellt。生物群落17:3(2011)314 - 316。
- 极化上皮细胞顶端膜的荧光显微镜- TIRF显微镜倒置。科学实验室(2011)//www.zns-mineralwater.com/science-lab/tirf-microscopy-of-the-apical-membrane-of-polarized-epithelial-cells/

